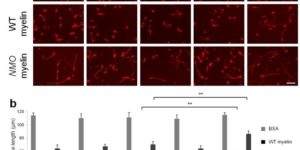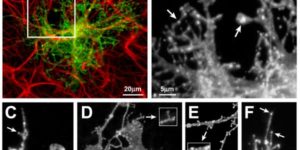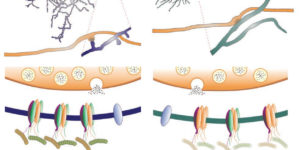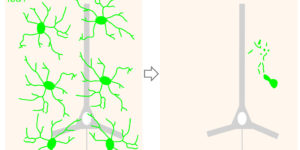
Hidden progenitors replace microglia in the adult brain.
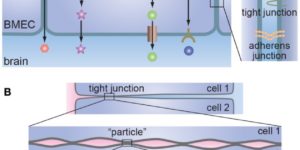
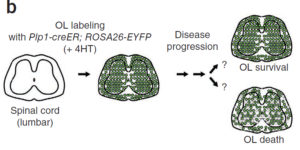
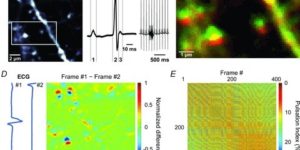
Microglia are highly dynamic components of the innate immune system. In this issue of Neuron, Elmore et al. (2014) report that global depletion of microglia triggers mobilization of latent microglial progenitors throughout the CNS, resulting in rapid repopulation.