Same players, different game: AMPA receptor regulation in oligodendrocyte progenitors.
Comment on “Bidirectional plasticity of calcium-permeable AMPA receptors in oligodendrocyte lineage cells.” [Nat Neurosci. 2011]
Comment on “Bidirectional plasticity of calcium-permeable AMPA receptors in oligodendrocyte lineage cells.” [Nat Neurosci. 2011]
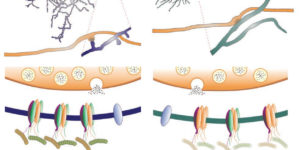
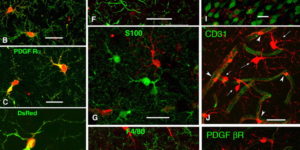
Oligodendrocyte precursor cells (OPCs) express NMDA receptors (NMDARs) and form synapses with glutamatergic neurons throughout the CNS. Although glutamate influences the proliferation and maturation of these progenitors in vitro, the role of NMDAR signaling in oligodendrogenesis and myelination in vivo is not known. Here, we investigated the consequences of genetically deleting the obligatory NMDAR subunit NR1 from OPCs and their oligodendrocyte progeny in the CNS of developing and mature mice. NMDAR-deficient OPCs proliferated normally, achieved appropriate densities in gray and white matter, and differentiated to form major white matter tracts without delay. OPCs also retained their characteristic physiological and morphological properties in the absence of NMDAR signaling and were able to form synapses with glutamatergic axons. However, expression of calcium-permeable AMPA receptors (AMPARs) was enhanced in NMDAR-deficient OPCs. These results suggest that NMDAR signaling is not used to control OPC development but to regulate AMPAR-dependent signaling with surrounding axons, pointing to additional functions for these ubiquitous glial cells.
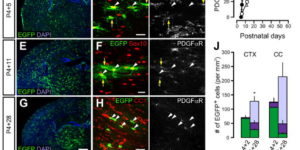
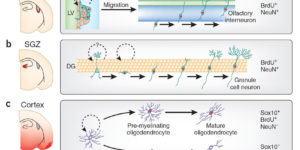
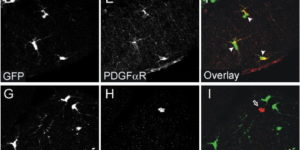
The mammalian CNS contains a ubiquitous population of glial progenitors known as NG2+ cells that have the ability to develop into oligodendrocytes and undergo dramatic changes in response to injury and demyelination. Although it has been reported that NG2+ cells are multipotent, their fate in health and disease remains controversial. Here, we generated PDGFαR-CreER transgenic mice and followed their fate in vivo in the developing and adult CNS. These studies revealed that NG2+ cells in the postnatal CNS generate myelinating oligodendrocytes, but not astrocytes or neurons. In regions of neurodegeneration in the spinal cord of ALS mice, NG2+ cells exhibited enhanced proliferation and accelerated differentiation into oligodendrocytes but remained committed to the oligodendrocyte lineage. These results indicate that NG2+ cells in the normal CNS are oligodendrocyte precursors with restricted lineage potential and that cell loss and gliosis are not sufficient to alter the lineage potential of these progenitors.
The ability to investigate the electrophysiological properties of individual cells in acute brain tissue led to the discovery that many glial cells have the capacity to respond rapidly to neuronal activity. In particular, a distinct class of neuroglial cells known as NG2 cells, which exhibit many of the properties that have been described for glial subtypes such as complex cells, polydendrocytes, synantocytes and GluR cells, express ionotropic receptors for glutamate and GABA. In both gray and white matter, NG2 cells form direct synaptic junctions with axons, which enable transient activation of these receptors. Electrophysiological analyses have shown that these neuron-glia synapses exhibit all the hallmarks of ‘classical’ neuron-neuron synapses, including rapid activation, quantized responses, facilitation and depression, and presynaptic inhibition. Electron microscopy indicates that axons form morphologically distinct junctions at discrete sites along processes of NG2 cells, suggesting that NG2 cells are an overt target of axonal projections. AMPA receptors expressed by NG2 cells exhibit varying degrees of Ca(2+) permeability, depending on the brain region and stage of development, and in white matter NG2 cells have also been shown to express functional NMDA receptors. Ca(2+) influx through AMPA receptors following repetitive stimulation can trigger long term potentiation of synaptic currents in NG2 cells. The expression of receptors with significant Ca(2+) permeability may increase the susceptibility of NG2 cells to excitotoxic injury. Future studies using transgenic mice in which expression of receptors can be manipulated selectively in NG2 cells have to define the functions of this enigmatic neuron-glia signaling in the normal and diseased CNS.
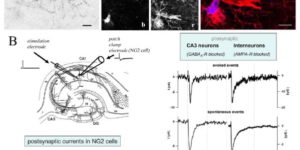
The mammalian CNS contains an abundant, widely distributed population of glial cells that serve as oligodendrocyte progenitors. It has been reported that these NG2-immunoreactive cells (NG2(+) cells) form synapses and generate action potentials, suggesting that neural-evoked excitation of these progenitors may regulate oligodendrogenesis. However, recent studies also suggest that NG2(+) cells are comprised of functionally distinct groups that differ in their ability to respond to neuronal activity, undergo differentiation, and experience injury following ischemia. To better define the physiological properties of NG2(+) cells, we used transgenic mice that allowed an unbiased sampling of this population and unambiguous identification of cells in discrete states of differentiation. Using acute brain slices prepared from developing and mature mice, we found that NG2(+) cells in diverse brain regions share a core set of physiological properties, including expression of voltage-gated Na(+) (NaV) channels and ionotropic glutamate receptors, and formation of synapses with glutamatergic neurons. Although small amplitude Na(+) spikes could be elicited in some NG2(+) cells during the first postnatal week, they were not capable of generating action potentials. Transition of these progenitors to the premyelinating stage was accompanied by the rapid removal of synaptic input, as well as downregulation of AMPA and NMDA receptors and NaV channels. Thus, prior reports of physiological heterogeneity among NG2(+) cells may reflect analysis of cells in later stages of maturation. These results suggest that NG2(+) cells are uniquely positioned within the oligodendrocyte lineage to monitor the firing patterns of surrounding neurons.
Comment on “PDGFRA/NG2 glia generate myelinating oligodendrocytes and piriform projection neurons in adult mice.” [Nat Neurosci. 2008]
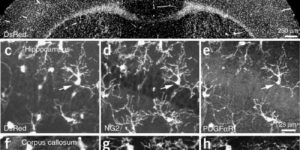
NG2 glia constitute a fourth major glial cell type in the mammalian central nervous system (CNS) that is distinct from other cell types. Although circumstantial evidence suggests that some NG2 glia differentiate into oligodendrocytes, their in vivo fate has not been directly examined. We have used the bacterial artificial chromosome (BAC) modification technique to generate transgenic mice that express DsRed or Cre specifically in NG2-expressing (NG2+) cells. In NG2DsRedBAC transgenic mice, DsRed was expressed specifically in NG2+ cells throughout the postnatal CNS. When the differentiation potential of NG2+ cells in vitro was examined using DsRed+NG2+ cells purified from perinatal transgenic brains, the majority of the cells either remained as NG2+ cells or differentiated into oligodendrocytes. In addition, DsRed+NG2+ cells also differentiated into astrocytes. The in vivo fate of NG2 glia was examined in mice that were double transgenic for NG2creBAC and the Cre reporter Z/EG. In the double transgenic mice, the Cre reporter EGFP was detected in myelinating oligodendrocytes and in a subpopulation of protoplasmic astrocytes in the gray matter of ventrolateral forebrain but not in fibrous astrocytes of white matter. These observations suggest that NG2+ cells are precursors of oligodendrocytes and some protoplasmic astrocytes in gray matter.
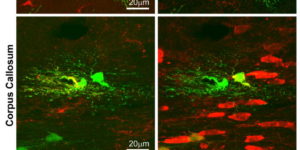
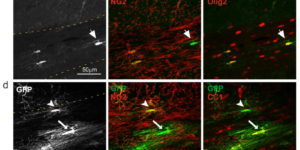
Directed fusion of transmitter-laden vesicles enables rapid intercellular signaling in the central nervous system and occurs at synapses within gray matter. Here we show that action potentials also induce the release of glutamate from axons in the corpus callosum, a white matter region responsible for interhemispheric communication. Callosal axonsreleaseglutamate by vesicular fusion, which induces quantal AMPA receptor-mediated currents in NG2(+) glial progenitors at anatomically distinct axo-glial synaptic junctions. Glutamaterelease from axons was facilitated by repetitive stimulation and could be inhibited through activation of metabotropic autoreceptors. Although NG2(+) cells form associations with nodes of Ranvier in white matter, measurements of conduction velocity indicated that unmyelinated fibers are responsible for glutamatergic signaling with NG2(+) glia. This activity-dependent secretion of glutamate was prevalent in the developing and mature mouse corpus callosum, indicating that axons within white matter both conduct action potentials and engage in rapid neuron-glia communication.
Chemical synaptic transmission provides the basis for much of the rapid signaling that occurs within neuronal networks. However, recent studies have provided compelling evidence that synapses are not used exclusively for communication between neurons. Physiological and anatomical studies indicate that a distinct class of glia known as NG2(+) cells also forms direct synaptic junctions with both glutamatergic and GABAergic neurons. Glutamatergic signaling can influence intracellular Ca(2+) levels in NG2(+) cells by activating Ca(2+) permeable AMPA receptors, and these inputs can be potentiated through high frequency stimulation. Although the significance of this highly differentiated form of communication remains to be established, these neuro-glia synapses might enable neurons to influence rapidly the behavior of this ubiquitous class of glial progenitors.
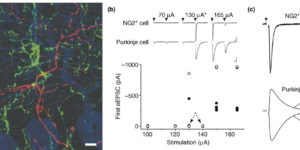
The molecular layer of the cerebellar cortex is populated by glial progenitors that express ionotropic glutamate receptors and extend numerous processes among Purkinje cell dendrites. Here, we show that release of glutamate from climbing fiber (CF) axons produces AMPA receptor currents with rapid kinetics in these NG2-immunoreactive glial cells (NG2+ cells) in cerebellar slices. NG2+ cells may receive up to 70 discrete inputs from one CF and, unlike mature Purkinje cells, are often innervated by multiple CFs. Paired Purkinje cell-NG2+ cell recordings show that one CF can innervate both cell types. CF boutons make direct synaptic junctions with NG2+ cell processes, indicating that this rapid neuron-glia signaling occurs at discrete sites rather than through ectopic release at CF-Purkinje cell synapses. This robust activation of Ca2+-permeable AMPA receptors in NG2+ cells expands the influence of the olivocerebellar projection to this abundant class of glial progenitors.